
Article by the Regenetarianism blog – all references linked below article
Reducing thermogenic carbon emissions and enhancing naturally occurring carbon sinks are both ways to balance the carbon cycle. Though instead of such both/and thinking, the choice often presented is an either/or one where either all thermogenic/biogenic emissions need to be quickly plus drastically reduced or the planet will go up in flames.
On the other hand, a few others seem to suggest that just enhancing sinks will somehow offset all thermogenic carbon emissions. Of course, this second point of view has led to a backlash from critics on the other side of this either/or equation.
These critics claim that sinks, particularly soil carbon sinks, are woefully inadequate to address and draw down all the excess carbon in the atmosphere. Though there’s some merit to some of their critiques, these critics too often have an outdated understanding of the ever-evolving soil science. So, some parts of their arguments are easy to quickly deconstruct.
Take for example, the 2022 opinion paper by Berthelin et al. (see link below) dismissing the potential of soil carbon to function as a significant carbon sink. These authors make their conclusion based upon what they believe is an overlooked phenomenon: the mineralisation of organic litter.
In a response letter to this opinion paper, Angers et al 2022 pointed out a few of the many shortcomings in Berthelin et al.’s logic including Berthelin et al.’s limited understanding of newer soil science paradigms.
To begin with, rapid mineralisation of organic inputs wasn’t some big new revelation or overlooked phenomena. But as Angers et al. pointed out, soil organic matter [SOM] isn’t only formed by decomposing litter piled on top of soil.
The MCP and mineral associated organic matter
As Angers et al detailed with its many cited references (e.g. Liang et al, 2019), another primary pathway for the formation of organic matter is via root exudates that feed microbes (the microbial carbon pump or MCP) that become necromass (microbial biosynthesis of carbon). That necromass, in turn, associates (binds) with minerals to become more recalcitrant forms of carbon including mineral associated organic matter or MAOM.
Most recent studies note that root carbon or exudates largely become MAOM (Angst et al 2024) whereas decomposing litter becomes particulate organic matter [POM]. POM isn’t as recalcitrant as MAOM, but POM can become occluded (blocked and trapped) in larger particles and last for decades if not disturbed (Mueller & Koegel-Knaber 2008, Angst et al 2023).
Respiration from or incorporation of added organic matter (litter) also depends on the existing microbiology of the soil organic matter upon which the new litter is being added. So, adding more litter to soils with higher amounts of existing soil organic carbon [SOC] incorporates more of the SOC from that new litter than adding litter to soils without much existing SOC. Why? The microbiological mix of the existing soil determines how much of the new soil carbon is integrated or released (Wu et al., 2024).
Decomposing fungi, in particular, in soils transform and move new litter into more recalcitrant forms of carbon further away from the detritusphere deeper into the soil profile. So, the mineralisation of all new matter doesn’t end up released into the atmosphere. Some of this litter is transformed and consumed by such decomposing forms of fungi. When this saprophytic fungi dies, its necromass also can associate with minerals to form MAOM (Witzgall et al., 2021). This, in part, is why soils with higher fungi to bacteria ratios sequester more and respire less carbon than soils that are more bacterial dominant (Wu et al., 2024).
Mycorrhizal pools of carbon
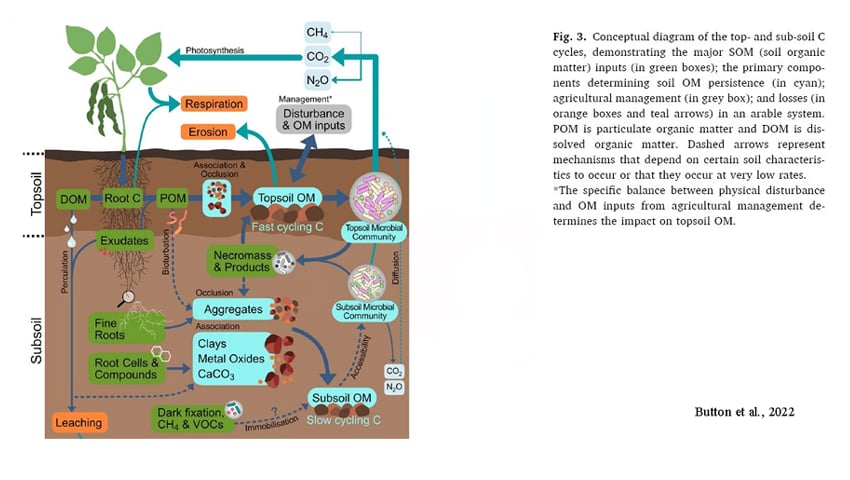
In addition to citing the MCP and decomposition pathways, Angers et al. noted that various agriculture practices (cover cropping, agroforestry, etc.) provide additional inputs of carbon into the soil via root exudates and from the roots themselves. These inputs aren’t so transient. Roots and the fungi that associate with or decompose them are transformed into forms of carbon (e.g. melanin, chitin) including necromass that last a lot longer than litter added to the top of soil (Button et al 2022). However Angers et al didn’t really get into how much other below ground biomass is retained in living/growing/dying mycorrhizae pools. These mycorrhizal pools of carbon for both ectomycorrhizae and endomycorrhizae are quite significant (Hawkins et al 2023).
Mycorrhizae also add a new wrinkle to the saturation debate that now revolves around the mineral content of soil for soil carbon to associate with. This wrinkle requires a little background. The old thinking is that it takes hundreds of year to form an inch of top soil via physical and chemical weathering. Per this thinking this is solely where minerals in soils come from and how “new” soil is formed.
Newer ideas on soil organic formation include the decomposition pathway which Berthelin et al recognises – though Berthelin et al argues that this litter provides very little carbon storage due to quick mineralisation of organic matter with less than 10% of that newly deposited organic matter becoming incorporated into soil carbon stocks. That is until soil carbon saturation is realized, and no more carbon can be added. These authors note that due to priming (carbon inputs adding to increased respiration by microbes), most carbon is respired, oxidized and re-emitted into the atmosphere. So their entire argument is that more and more organic matter has to be piled onto the top of soil to get anywhere near close to having any significant impact on reducing atmospheric levels of carbon. Thus Berthelin et al simply conclude there’s not enough organic matter that can be added to the top of soil to achieve any meaningful reduction of atmospheric carbon.
Minerals and mycorrhizae
Now newer thinking on soil organic matter (SOM) formation includes the MCP (mentioned above) in addition to the decomposition pathway. Per this newer thinking, SOM is divided into two different types of SOM based on fraction size.
Larger matter is particular organic matter or POM. This is still constrained by issues of mineralisation and respiration as detailed by Berthelin et al. Smaller denser organic matter is mineral associated organic matter or MAOM. Though there’s some debate, the current prevailing belief is that there’s no limit on POM formation but there is an upward limit on MAOM formation (Cotrufo et al 2019).
What limits MAOM per this thinking is the amount of minerals in soil for soil organic carbon (from either the decomposition pathway or MCP) to attach to (i.e. associate with) and form MAOM. These minerals, per this train of thought, were solely made available from bedrock through physical and chemical weathering that takes hundreds of years to occur. Thus the newer thoughts on soil carbon saturation revolve around the limitations of minerals to associate with and the longer timeframe it would take to weather more minerals. Therefore, once all the minerals are used up to associate with, soils again “saturate” and no more carbon can be stored.
But even people, who ascribe to this theory, believe that current saturation of top soil and subsoil are respectively around 42% and 21% (Georgiou et al 2022). So there’s still plenty of minerals to associate with to draw down atmospheric carbon in many places.
This is where mycorrhizae come in and need to be considered.
Mycorrhizal fungi source macro-nutrients, micro-nutrients and water for plants by drastically increasing the root zones of plants into subsoil all the way down to bedrock in some places. The hyphae in this mycorrhizosphere exude acids that weatherise minerals and make them plant available.
Thus in addition to providing plants nutrients, mycorrhizae also increase mineral availability in soils that carbon can associate with to form more recalcitrant forms of carbon namely MAOM (Wu et al 2023). This is referred to as the soil mineral carbon pump or MnCP (Xiao et al 2023).
Many of the micro-nutrients mycorrhizae provide are also essential for photosynthesis. For example manganese is needed for the hydrolysis of water which provides the free electrons needed for the Calvin Cycle. Thus photosynthetic rates can be increased by correcting for any micro-nutrient deficiencies. Greater rates of photosynthesis means more root exudates can be pumped into the soil via the MCP.
Measuring soil carbon stocks
There’s another problem with Berthelin et al.’s saturation argument. This problem revolves around how soil carbon stocks are determined.
The vast majority of measurements are based on the “fixed depth method” to depths of 20 or 30 centimeters.
This shallow depth is largely chosen for speed, cost and ease (Yost et al 2020) plus the understanding that this shallow depth is what’s most impacted by management changes (FAO 2022).
However, taking measurements over the course of time using this fixed depth method doesn’t properly account for changes in soil density, accrued organic matter, and MAOM formed deeper in the soil profile.
Since soil organic stocks are based on the multiplication of soil organic matter, depth and soil bulk density, reliance on fixed depth measurements is very problematic and potentially inaccurate (Fowler et al 2023, Raffeld et al 2024).
So, reliance on this fixed depth method to shallow depths, based on limitations of the maths, may give the false impression that soil sinks are becoming “saturated” because they’re not accounting for how increasing soil carbon actually decreases soil bulk density by improving soil structure allowing for greater air and water permeability.
Thus, many aren’t recognising that the size of soil sinks isn’t finite. This seems to be the case with many researchers (Smith 2014, Wang et al 2023) doing soil carbon modeling using available data but not actually doing any of their own field measurements.
Moreover, using these shallow metrics also doesn’t account for more resilient forms of soil organic matter that are getting buried including occluded POM and especially MAOM (Button et al, 2022, Angst et al 2023, Mossier et al 2023).
Ideally soil carbon would be measured to the depth of root tips. For perennial systems, this could be quite deep. So, using a depth of at least 60 to 100 centimeters is more realistic as well as other accounting methods like equivalent soil mass or ESM (Raffeld et al 2024).
Water and soil inorganic carbon
Additionally, Berthelin et al.’s assumption that all respired carbon by microbes oxidises and returns to the atmosphere isn’t entirely correct especially in semi-arid and arid high pH soils.
Here, where water gets temporarily retained in the soil, respired CO2 reacts with that water to form carbolic acid.
This carbolic acid reacts with minerals in the soil like calcium to form calcium carbonate (Naorem et al 2022). Calcium carbonate is a form of soil inorganic carbon or SIC. (Inorganic carbon as defined here is a simple carbon molecule without two carbon atoms bound together by a double bond).
So, seeing that approximately 40% of the earth’s soil is in such semi-arid and arid places, there’s also a huge pool of soil inorganic carbon that largely forms in the deeper B horizons that also isn’t being accounted for in soil organic carbon models and inventories.
Though there’s not as much research emphasis on SIC as there is on SOC (Raza et al. 2024) largely because most soil carbon testing has been done in the Northern Hemisphere. Soil carbon stock maps also use the Mercator Map projection which makes continents (and consequently SOC stocks) in the Northern Hemisphere look much larger than those in the Southern Hemisphere.
Note too in soil ecosystems that have been recarbonated, due to improved soil structure allowing for better water utilisation and increase microbial diversity, more above and below ground biomass is possible. These are both large carbon pools.
Plus, when ground is covered with such above ground biomass (plants), carbon respired by microbes is also a large source of CO2 for these plants to use for photosynthesis. It’s not a coincidence that stomata are located on the underside of a plant’s leaves. So respired carbon is also constantly being recycled where there is continuous ground cover (Brinton 2015).
Anyway, as can be seen from how recent many of the soil science references cited above are, soil science is constantly evolving.
So, being dogmatic about any of this science is really a folly.
But nonetheless, many scientists tend to get trapped in their outdated paradigms. So, accepting newer scientific paradigms is not something they are wont to do.
References:
Angst et al 2023 Unlocking complex soil systems as carbon sinks: multi-pool management as the key
Brinton, Will 2015. Farming the CO2 Factor
Georgiou, K et al 2022 Global stocks and capacity of mineral-associated soil organic carbon
Hawkins, H-J. et al Mycorrhizal mycelium as a global carbon pool
Smith, P 2014 Do grasslands act as perpetual carbon sinks
Wang, Y. et al 2023 Risk to rely on soil carbon sequestration to offset global ruminant emissions
Xiao, K-Q. et al 2023 Introducing the soil mineral carbon pump